МОДЕЛЬ ЭВОЛЮЦИИ ЭТНОСОВ И ЕЕ РЕАЛИЗАЦИЯ НА БАЗЕ ГЕНЕТИЧЕСКИХ АЛГОРИТМОВ
(г. Камышин Волгоградской обл.)
Предлагается математическая модель,
отражающая основные этапы зарождения, развития и угасания этносов на основе
представлений, изложенных в работах Л.Н. Гумилева. В модели, воспроизводящей фазы этногенеза, содержатся элементы, имитирующие
влияние биологических факторов («мотор» развития этноса), особенностей
ландшафта (воздействие внешней среды) и истории (идентификация модели).
Биологический фактор моделируется с использованием генетических алгоритмов.
Приводятся результаты моделирования для различных сочетаний факторов, влияющих
на изменение пассионарности этноса. Излагаются перспективы развития и уточнения
модели.
THE SIMULATION
MODEL OF ETHNOS EVOLUTION AND ITS IMPLEMENTATION ON THE BASIS OF GENETIC
ALGORITHMS
Krushel E.G.,
Surgutanov V.V.
(The city of Kamyshin,
Volgograd region)
The
issue deals with the mathematical model for the simulation of the main stages
of ethnos appearance, arising, stagnation, and destruction on the basis of L.N.
Gumilev’s ideas. The model reflecting the phases of ethnogeny consists of the
elements for simulation of the biology factors (the evolution driver), the
landscape features (the external environment influence) and history (model
identification). The biology factor is simulated by means of genetic algorithms
application. The simulation results are presented for the different combination
of the factors effecting the ethnos
energetic capacity. Some perspectives of model improvement are stated.
Известно, что исторические модели
сложны для построения, поскольку необратимость времени ведет к отсутствию
массовых объектов для идентификации статистических характеристик и, как
следствие, для обоснованного прогнозирования. Поэтому представляет интерес
построение формальных моделей для генерации возможных сценариев исторического
процесса на основе некоторых гипотез, предлагаемых (умозрительно) в трудах
историков и философов. Одним из вариантов
является подход, позволяющий моделировать зарождение, развитие и гибель
абстрактного этноса на базе формализации гипотез, предложенных в работах
историка Л.Н. Гумилева [1]. Согласно данным гипотезам этногенетический процесс
проходит определенные фазы, характеризующиеся различным уровнем суммарной
энергетики этноса, именуемой пассионарностью.
Основные фазы: инкубационный период, пассионарный подъем, ведущий к
акматической фазе, фаза надлома и фаза инерции, после чего непременно
наблюдаются фазы обскурации и гомеостаза. Высокий уровень пассионарности
свидетельствует о силе этноса, о его способности адаптироваться и сопротивляться
действию окружающей среды. Низкий – о малом количестве энергичных особей, по
сравнению с общим числом членов этноса. Этногенез трактуется Л.Н. Гумилевым как
синтез трех дисциплин: биологии; географии; истории.
Согласно трем компонентам удается построить модель, воспроизводящую похожие фазы этногенеза и имитирующую биологические факторы как мотор эволюции, особенности ландшафта как воздействие внешней среды и ресурсные ограничения и историю для идентификации модели.
Назначение модели эволюции этноса. Моделирование этногенеза имеет определенный теоретический интерес. В большинстве работ, посвященных созданию моделей этногенеза [2], постулируется правильность кривой изменения пассионарности этноса во времени, предложенной Л.Н. Гумилевым, а затем предлагаются уравнения, решение которых позволяет получить похожую зависимость. В отличие от этого в данной работе предлагается модель с элементами саморазвития, подчиняющегося эволюционным законам и ограничениям. Если окажется, что такая модель «самостоятельно», без вмешательства процессов идентификации параметров воспроизведет фазы изменения пассионарности этноса, то это послужит не только обоснованием самой модели, но и даст косвенное подтверждение теории.
Высокий уровень абстракции и стохастичность предметной
области, в которой протекает исторический процесс, ведет к невозможности
отыскания прямых доказательств или опровержений теории. Дискуссии об
адекватности выдвинутых гипотез часто
используют аргументы, основанные лишь на единичных фактах. Недостатком же
исторических фактов является их неполная объективность. Подход, основанный на
математическом моделировании, дает уверенность в том, что имитируемые в ней
факты будут воспроизводимыми. Такое
положение вещей связано с тем, что любой результат, полученный при
моделировании, может быть проанализирован; при этом всегда удастся выявить
причину, обусловившую данный результат. Процесс отыскания истоков тех или иных
явлений, демонстрируемых в модели, может быть сколь угодно сложен, но
однозначен. Поэтому моделирование динамических процессов является очень
продуктивным, позволяя находить пусть косвенные, но объективные аргументы в
пользу принятия или непринятия теоретических положений.
Кроме того, моделирование выгодно в системах с большим числом случайных факторов. Модель устраняет несущественные из них и является приближением реальной динамической системы к автомодельной системе.
При рассмотрении пассионарной теории этногенеза
легко увидеть, что параметром, являющимся движущей силой автомодельного
этногенетического процесса, служит пассионарность. Если избрать его стержнем модели, то появляется надежда вскрыть
структуру и параметры динамики исторического процесса.
Для понимания сути исследования необходимо
подробнее изложить основные положения, принятые при алгоритмизации. Наиболее
простая модель предполагает наличие изолированного этноса. Этнос как объект
исследования представляется в виде совокупности пассионарных и субпассионарных
особей, проживающих в определенном ландшафте. На пассионарность особей оказывают
влияние наследственность, изменчивость и естественный отбор, обусловленный
системой ресурсных и иных ограничений. Под иными подразумеваются ограничения,
вызванные действием посторонних сил, внешних, не обусловленных главным
автомодельным параметром (пассионарностью этноса). Таким образом, в модели
удается неявно учесть влияние соседних этносов.
Будучи сильно упрощенной, такая модель позволяет анализировать адекватность лишь той части гипотез, которые касаются внутренних свойств этноса. Основным из них является наличие фаз пассионарного напряжения. Менее изученным, но не менее показательным является свойство нестабильности этноса в моменты перехода в следующую фазу.
Алгоритмическое обеспечение. Инструментом моделирования избраны генетические алгоритмы [3]. Такой выбор обусловлен действием эволюционных и инволюционных закономерностей, управляющих развитием этносов и похожих на законы естественного отбора (формализация которых и привела к появлению генетических алгоритмов). Составляющими генетического алгоритма являются механизмы мутаций, наследственности и естественного отбора.
Генетический алгоритм реализуется
следующими действиями:
1.Создание начального числа особей (объектов) в количестве n.
2.Наделение объектов хромосомами, содержащими значения генов, заданных
случайным образом либо детерминированных.
3.Случайный выбор пары объектов. Вероятность выбора объекта прямо пропорциональна величине критерия «привлекательности» особи, приходящейся на единицу ресурса.
4.Создание пары потомков, наследующих гены объектов родителей согласно случайно осуществленному делению хромосом.
5.Генерация случайного события (возникновения мутации) в соответствии с заданным законом распределения вероятностей появления мутаций.
6.В случае, если будет мутация, случайным образом выбирается хромосома и мутирующий ген объекта-потомка. Затем случайным образом задается значение мутирующего гена.
7.В популяцию добавляются два созданных объекта – потомка.
8.В случае, если ресурсные ограничения выполнены, из популяции удаляются два объекта, имеющих наименьшее значение приращения критерия на единицу ресурса.
9.Если же ресурсные ограничения не выполнены, то, возникает эффект недоверия, и из популяции удаляется пара особей потомков, добавленная в п. 7.
10.Алгоритм возвращается к п.3. Завершение алгоритма происходит тогда, когда общесистемный критерий эффективности популяции перестает улучшаться.
Сходимость генетических
алгоритмов не доказана, но все попытки их использования привели к положительным
результатам.
Применение генетических
алгоритмов при моделировании эволюции этносов. В описанной форме генетические алгоритмы
используют в задачах оптимизации, не имеющих генетическую природу. Процессы этногенеза имеют генетическую
подоплеку. Их результат диктуется эволюционными законами развития.
Использование генетических алгоритмов для формализации этногенетических
процессов имеет целью не оптимизацию, а обоснованное моделирование. Часть
генетического алгоритма (шаги которого описаны выше), отвечающая за
естественный отбор, не является прямым следствием эволюционной теории
организмов и используется как средство управления процессом эволюции объектов.
В этом отличие использования генетических алгоритмов для оптимизации от их
использования в качестве инструмента моделирования.
|
Рис. 1. |

В рамках пассионарной теории этногенеза
использование генетических алгоритмов происходит при следующих допущениях:
1.Каждую особь s, входящую в этнос, описывают вектором ps в пространстве, координатами которого являются всевозможные виды деятельности человека, именуемые призваниями (рис.1). Для реализации призвания требуются энергетические затраты, черпаемые особью из ландшафта. Проекции этого вектора (ps)i на координатные оси есть энергия, расходуемая особью на совершение работы в направлении Ai, определяемом осью.
2.Ландшафт, вмещающий этнос, обладает конечным суммарным запасом энергии E,
который может быть потреблен этносом.
3.В энергетическом пространстве определен единичный вектор гармонии H. В начале процесса этногенеза энергетические вектора особей ps совпадают с вектором гармонии H. Именно такая длина и направление вектора позволяет особи обеспечивать жизнедеятельность при отсутствии необходимости к адаптации.
4.Процессы мутации, естественного отбора и наследования признаков есть процессы стохастические.
5.Благодаря мутациям в этносе возникают особи, обладающие избыточной
энергией и именуемые (по Л.Н. Гумилеву) пассионариями
(проекция ps на H>1), а также особи, обладающие недостатком
энергии – субпассионарии (проекция ps на H<1).
Вероятность мутации P0 пропорциональна скорости мутаций V.
6.В отсутствие мутаций вектор призваний объекта-потомка есть комбинация элементов векторов призваний объектов-предшественников pki = psi,, где k – индекс потомка, s – индекс одного из предшественников, i Î I.
7.Вероятность P1 участия особи в процессе воспроизводства
объектов-потомков прямо пропорциональна энергии особи |ps| (P1~|ps|). Данный принцип
является следствием привлекательности пассионариев,
а также следствием эффекта индукции.
8.Естественный отбор способствует выживанию особей, отклонение вектора призваний ps которых от вектора гармонии H минимально. Естественный отбор ужесточается при усилении действия внешней среды 1/K, где К- лояльность среды . Обозначив вероятность гибели особи P2 и учитывая независимость событий, определим действие естественного отбора так: P~|ps-H|/К.
9.Уровень пассионарности L есть скаляр, равный сумме проекций ps на H.
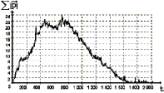
Некоторые результаты моделирования. Программный комплекс, разработанный в соответствии с описанными положениями, позволяет моделировать изменение следующих характеристик этноса во времени: уровней пассионарности и энергии этноса или отдельной особи, а также числа особей в этносе. Параметрами, варьируемыми при моделировании, являются: начальное число особей этноса, количество призваний, свойства (энергия) ландшафта, вероятность мутаций, продолжительность периода эволюции этноса. Типичный график изменения пассионарности абстрактного этноса представлен на рис. 2. Полученный результат является косвенным доказательством гипотезы о наличии фаз этногенеза. Таким образом, модель позволяет иллюстрировать самый общий, лежащий на поверхности принцип динамики исторического процесса.
|
Рис. 2. |
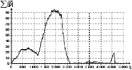
Другим результатом является иллюстрация утверждения о том, что с течением времени этнос ослабевает настолько, что становится уязвимым даже для самых слабых соседей, окружающих его. Данный результат получается при искусственном варьировании параметра “лояльности среды”. Как видно из графика, рост пассионарности возможен даже в стадии надлома, при слабых соседях. На стадии же гомеостаза даже самые лояльные соседи не позволяют этносу проявлять уже угасшую пассионарность (рис. 3).
|
Рис. 3. |
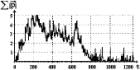
В числе прочих, модель иллюстрирует и свойство нестабильности этноса в моменты перехода от одной фазы этногенеза к другой.
Полученные результаты, косвенно подтверждающие гипотезы Л.Н. Гумилева, подлежат тщательному анализу и выявлению причин, обусловивших такое поведение модели. На данном этапе необходимо отметить, что уменьшение уровня пассионарности этноса связано, в основном, с уменьшением числа степеней свободы генома, в связи с адаптацией к условиям ландшафта.
|
Рис. 4. |
Сходство результатов
моделирования с реальными историческими процессами позволяет с большим доверием
относиться к перспективам практического использования теории. В данном
контексте модель превращается в инструмент, позволяющий учиться на ошибках, не
совершая их в реалии. Так, генерируя множество альтернатив, варьируя параметры
имитационной модели, можно получить ряд полезных с практической точки зрения
выводов.
Одним из них является принцип “национальной идеи”. В эксперименте такая ситуация моделируется изменением количества призваний. Для первого графика (рис. 4) их 3 для второго (рис. 5) – 5. Видна сильно нелинейная зависимость уровня пассионарности от количества призваний, при прочих равных условиях.
|
Рис. 5. |

Другим важным результатом моделирования является вскрытие причин, влияющих на продолжительность жизни этноса, например, для этноса, обеспечивающего высокий уровень пассионарности за счет привлечения ресурсов других стран (в том числе и интеллектуальных ресурсов); для этноса, имеющего высокий процент городского населения; для недальновидного этноса, экспортирующего сырье и др.
Перспективы повышения уровня адекватности модели. Недостатком описанной упрощенной модели является
то, что она не учитывает многие феномены исторического, этногенетического
процесса. Ведь многообразие форм его проявления во многом имеет место благодаря
многогранному процессу адаптации и взаимодействия этноса не только с
ландшафтами, но и с другими этносами, их населяющими. Создание такой модели
позволило бы имитировать множество фактов и особенностей глобального, мирового
этногенеза, а также многие важные, в том числе и онтологические аспекты
этногенетического процесса, до сих пор являющиеся белыми пятнами пассионарной
теории этногенеза.
Основным предметом спора является вопрос о причинах зарождения этноса. Как тело этноса в фазе гомеостаза может служить почвой для появления новой жизни – этноса инкубационного периода? Почему этнос зарождается лишь на территориях, являющихся пограничными для ландшафта? Чем определяются строгие временные ограничения срока жизни этноса?
Попыткой ответить на эти до сих пор нерешенные вопросы является модель, учитывающая взаимное влияние этносов посредством этнических контактов в форме симбиоза, ксении и химеры [4]. Кроме того, модель должна содержать ландшафты различных типов, с разной степенью сложности адаптации к ним. Это требование позволит получить число степеней свободы, достаточное для поддержания сложности и разнообразия в глобальной этногенетической системе человечества.
Ответ на нерешенные вопросы может дать механизм, в полной мере присущий популяционной генетике, т.е. механизм адаптации. Известно, что адаптация одновременно является и ускорителем эволюции, и ее тормозом. Человек является существом, наделенным поразительными способностями к адаптации по сравнению с другими живыми существами. Данное качество позволяет ему не только приспосабливаться к условиям внешней среды, но и адаптировать ее под свой режим существования. Если провести параллель между организмом и этносом, то можно прийти к ряду выводов, позволяющих надеяться на обнаружение причин зарождения этноса.
|
Рис. 6. |

Итак, отождествление этноса организму ведет к рассмотрению его сквозь призму процессов адаптации. Для этноса внешней средой являются окружающие его этносы, принадлежащие либо к общему, либо к разным суперэтносам. Принадлежность к тому или иному суперэтносу восходит к истокам исторического развития человечества, при этом не исключено и влияние различия в ландшафтах. Кроме того, внешней средой для этноса служит ландшафт, его вмещающий (рис. 6). Каждая из составляющих окружения этноса требует строго определенного направления адаптации (приспособления к себе). Обладая собственным, присущим только ему направлением вектора пассионарности, этнос вовлекается в процедуру приспособления к ландшафту и другим этносам, адаптируясь и адаптируя внешнюю среду. Очевидно, что способности организма к адаптации ограничены и, проводя параллель с организмами, приходим к теории эволюции этносов, аналогичной теории суровых ландшафтов [5]. Основным положением данной теории является принцип, согласно которому появление жизнеспособных особей в слишком жестких условиях маловероятно. Поэтому возникновение и развитие этноса вблизи другого развивающегося этноса, принадлежащего иному суперэтносу, является маловероятным. Тяжесть резкого, контрастного приспособления нашла отражение и в других генетических теориях, например в законе Боннера о низкой плейотропии [5].
Рассматривая лишь эту грань призмы адаптации, в итоге придется признать, что такое положение вещей привело бы к тому, что более древний этнос, имея больше времени на постепенную изменчивость своего генофонда пассионарности, в итоге сгенерирует суммарное направление своего вектора призваний, максимально адаптированное к ландшафту. При этом такой гигант легко расправится с любыми нарастающими на его теле молодыми, малочисленными этносами, которые вынуждены бороться не только с “супердержавой”, но еще и с враждебным для себя ландшафтом. Из всего этого последовала бы невозможность этногенеза, а, следовательно, неправомерность выдвинутой гипотезы о сходстве организма и этноса.
Но в данном рассмотрении не учтен один
немаловажный и достаточно новый факт из области современной генетической науки.
Адаптация - это верный путь к деградации [5]. Максимально адаптированный
организм теряет способности к изменчивости и становится уязвимым для
конкурентов по естественному отбору. Так, абсолютно адаптированный организм
обладает нулевой степенью свободы своего генома. Тем самым он отождествляется
этносу, находящемуся в состоянии гомеостаза. Таким образом, малейшее химерное
образование разрушит его скелет, внешне внушительный, но внутренне крайне
хрупкий. Именно данные две грани адаптационного механизма обеспечивают
некоторый оптимальный уровень развития этноса, ведущий к постоянной конкуренции
и поддержанию сложности структуры исторического общества.
Эти принципы являются основой функционирования не только генетических образований, но и систем любой произвольной природы. В области сложных систем замечено, что долговечность и живучесть системы зависит от поддержания необходимого уровня сложности ее внутренней структуры.
Список литературы:
1.Л.Н. Гумилев, "Этногенез и биосфера Земли", М, 1993.
2.http://gumilevica.kulichki.net
3.http://www.neuroproject.ru/genealg.htm
4.http://kraevedenie.chat.ru/sam7.htm
5.http://saisa.chat.ru/ga/a-z/a.htm.