КИНЕТИКА КЛЕТОЧНЫХ ПОПУЛЯЦИЙ
В ЭКОТОКСИКОЛОГИЧЕСКИХ МОДЕЛЯХ
Ершов Ю.А., Котин В.В.
(Москва)
Детерминированные
и стохастические задачи динамики популяций неоднократно рассматривались ранее [1,
3, 4] и традиционны для математической экологии. Вместе с тем интерес к этим
задачам, особенно стохастическим, постоянно возобновляется [5-7]. В настоящей
работе исследуются экотоксикологические модели, описывающие воздействие
химических агентов на рост клеток [12, 13].
1. Детерминированная экотоксикологическая
модель.
Уравнения
экотоксикологической детерминированной модели в нормированном виде записываются
следующим образом [12]:
 (1)
(1)
где
 (2)
(2)
Здесь z1, z2
– населенности двух групп клеток, нормированные на емкость b/a среды без
воздействия токсикантов: z1=c1a/b, z2=cma/b,
t=pt – безразмерное время,
c1, cm
– населенности групп растущих и митотических клеток; a,b,p -кинетические
коэффициенты автоингибирования, рождения (разветвления), и роста популяционной
цепи, w1 – мощность внешнего источника клеток с1, f –
коэффициент размножения. Коэффициенты b1 и p1 являются
функциями от концентрации ингибиторов x1 и x2:
px=p+d1;
bx = b + d2 , d1 = d11 x1
+ d12 x2; d2 = d21 x1 +
d22 x2 .
Известно, что разделение
характерных временных масштабов в задачах биокинетики позволяет упростить
анализ и выявить существенные особенности взаимосвязанных процессов [3].
Соотношение характерных времен двухстадийного роста клеток первой z1
и второй z2 группы зависит от параметра m=b/p. Если константы скорости
рождения b и продолжения цепи p соотносятся таким образом, что m>>1, то полная система (1)
редуцируется к вырожденному (медленному) уравнению:
![]() , (3)
, (3)
где
![]()
Эффект ингибирования –
уменьшение стационарного значения населенности zst -эквивалентен
снижению эффективной емкости среды при наличии токсикантов. Найдем диапазон
концентраций токсикантов, при которых стационарная населенность двухуровневой
популяции zst(х1,х2) не меньше заданной
величины sі0. Множество значений концентраций токсикантов {х1,х2},
удовлетворяющих этому условию, определяется неравенством
![]() (4)
(4)
где
![]()
В плоскости (х1,х2)
условие (4) задает область {0Јx1Јx1*, 0Јx2ЈF(x1,s)}, в пределах которой концентрация токсикантов
такова, что рост популяции возможен. Нулевому уровню стационарной населенности
s=0 в плоскости параметров (х1,х2) соответствует критическая кривая биоцидного действия zst(х1,х2)=0,
задающая верхнюю границу области роста G. Если концентрации токсикантов
превышают критический уровень, определяемый границей s=0, то рост популяции
полностью подавляется. Рис.1 демонстрирует зависимость стационарной
населенности zst(х1,х2) от концентрации
токсикантов для популяции без источников (W=0). Кривые постоянного уровня
населенности и критическая кривая биоцидного действия zst(х1,х2)=0
показаны на рис.2а.
В системе без источников
(W=0) нулевой уровень zst(х1,х2)=0 является
бифуркационной кривой, на которой перестраивается фазовый портрет. Критическая
кривая биоцидного действия ВС разбивает плоскость параметров (х1,х2)
на области G и R (рис. 2а). В области G плоскости параметров (х1,х2)
существует две особых точки – седло (0,0) и устойчивый узел (z*1,z*2).
В области R плоскости параметров (х1,х2) существует
единственная особая точка – устойчивый узел (0,0). На критической кривой ВС
устойчивый узел (z*1,z*2) сливается с седлом (0,0),
образуя множественную особую точку седло-узел, которая при дальнейшем росте
концентрации токсикантов и переходе в область R трансформируется в устойчивый
узел. Фазовые портреты системы в областях R и G пространства параметров (х1,х2)
показаны на Рис. 2б. Математический анализ детерминированной
экотоксикологической модели и графическое представление фазовых траекторий
реализованы с использованием пакета MAPLE V R5 [14].

Рис. 1. Стационарная
населенность zst в зависимости от концентрации токсикантов для
изолированной популяции (при отсутствии внешнего потока клеточной массы W=0). Показаны
кривые постоянного уровня стационарной численности популяции zst(х1,х2)=const.
Нулевой уровень zst(х1,х2)=0 соответствует
критической кривой биоцидного действия.

а) б)
Рис.2. (а) Бифуркация на
критической кривой биоцидного действия BС. В области G две особых точки – седло
(0,0) и устойчивый узел (zst1, zst2).
В области R единственная особая точка- устойчивый узел. На BС – множественная особая
точка седло-узел (0,0). (б) Фазовые портреты системы в области R при х1=2,
х2=5, p/b=0.1 и в области G, p/b=0.02.
2. Стохастическая динамика клеточных популяций и время жизни.
Клеточную популяцию в среде
со случайными возмущениями рассмотрим как нелинейную динамическую систему с
флуктуациями [2, 5-7]. Приемлемой моделью быстрых флуктуаций обычно является
стационарный дельта-коррелированный случайный процесс (белый шум). Выделим два
типа возмущений. Первый тип- аддитивные шумы (флуктуации мощности внешнего
источника dW). Второй тип–
параметрические шумы (случайные изменения кинетических коэффициентов),
приводящие к появлению в уравнениях стохастической динамики мультипликативных
членов, зависящих от состояния системы в данный момент времени [5].
Математическое ожидание
времени первого достижения заданных границ zmin, zmax
находится из решения краевой задачи [2, 6, 13]:
![]() , zmin<z<zmax (5)
, zmin<z<zmax (5)
T(zmin)=0; T(zmax)=0, a(z) и b(z)
– коэффициенты сноса и диффузии в уравнении Фоккера-Планка-Колмогорова.
Выбор нижней границы zmin=0
(или zmin=zВ, где zВ–нижний порог численности,
по достижении которого популяция вырождается) соответствует условию гибели
популяции. Верхняя граница zmax задается исходя из ограничений
ресурсов среды.
Интегральное описание
вероятностных темпоральных
характеристик системы, эволюционирующей в случайной среде, подразумевает
усреднение функции T(z) по начальному распределению численности r(z)
![]() (6)
(6)
Такое усреднение учитывает
зависимость времени жизни от формы и дисперсии начального распределения r(z),
а также от степени близости начального состояния (хвостов начального
распределения) к границам жизнеспособности популяции zmin, zmax
[13].
Итак, <T> – среднее
время существования популяции в пределах ограниченной численности z Н[zmin,zmax], рассматривается как интегральная
вероятностная характеристика, определяющая усредненный временной интервал существования
популяции в пределах ограниченной численности.
3. Влияние флуктуаций и токсикантов на время
жизни популяции.
Исследуем динамику
экотоксикологической системы в случайной среде, используя для редуцированной
модели аналитический подход, а для анализа двухстадийного роста ингибированной
клеточной популяции метод статистического моделирования. В качестве допустимой
области существования популяции выделим ограниченный интервал [zmin,zmax].
Случайные возмущения приводят к тому, что изображающая точка M(z) в некоторый случайный момент времени достигает
граничных значений zmin, zmax, которые в соответствии с
введенным выше определением интерпретируются как допустимые пределы
населенности популяции. На Рис. 4,5 представлены результаты численного решения
двухточечной краевой задачи (5). При расчетах варьировались координаты
граничных точек zmin, zmax, фактор
ингибирования l (токсическое воздействие на
популяцию) и среднеквадратичная интенсивность случайных возмущений s.
Оказывается, что форма начальных распределений весьма незначительно влияет на
усредненное время жизни <T>. Основным параметром начального
распределения, влияющим на величину <T>, является дисперсия
распределения, отнесенная к ширине интервала между границами.
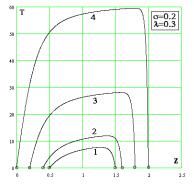

|
Рис. 4. Время достижения
заданного уровня населенности популяции (s=0.2; l=0.3) для различных границ по населенности
популяции: 1) zmin=0.5; zmax=1.5; 2) zmin=0.4;
zmax=1.6; 3)zmin=0.2; zmax=1.8; 4) zmin=0;
zmax=2. |
Рис. 5. Время первого
достижения заданного уровня населенности популяции для различных значений
фактора ингибирования l; zmin=0; zmax=2. |

Рис. 6. Случайное
блуждание на фазовой плоскости (z1,z2)
под действием флуктуаций токсичности.
В процессе статистического моделирования
двухстадийного роста ингибированной клеточной популяции наряду с аддитивными
(внешними) случайными возмущениями в систему вводились также параметрические
возмущения, соответствующие вариациям концентрации токсиканта и коэффициента
скорости роста клеток. Случайные возмущения моделировались в среде Mathcad 8
[15] посредством различных вариантов программных датчиков случайных чисел
(число случайных испытаний N~104 -105).
На Рис. 6 показана
реализация случайного процесса блуждания изображающей точки на фазовой
плоскости (z1,z2) под действием флуктуаций токсичности.
Результаты моделирования совместного действия аддитивного и параметрического
шума для двух значений мощности внешнего источника W=1, W=2 иллюстрируются на
Рис. 7.

Рис. 7. Случайное блуждание
на фазовой плоскости (z1,z2) при совместном действии
флуктуаций токсичности и внешнего источника для двух значений средней величины
потока клеточной массы (W=1; W=2).
Выводы.
Действие аддитивных и
параметрических шумов делает токсическую границу биоцидного действия в
экотоксикологической модели «проницаемой» для нелинейного броуновского
блуждания [7] изображающих точек в фазовом пространстве. Существует отличная от
нуля вероятность выхода изображающих точек за пределы граничных состояний, даже
если детерминированные параметры системы, обеспечивают рост популяции. С другой
стороны, если детерминированный рост клеточной массы подавлен токсикантами, но
система находится в окрестности критической кривой биоцидного действия, то
существует вероятность роста клеточной массы.
Модель описывает
популяционную динамику экосистем в условиях токсического действия загрязнений
окружающей среды, а также характеристики опухолевого роста с учетом совместного
действия цитостатиков, канцерогенов и случайных возмущений.
Литература.
1.
Вольтерра
В. Математическая теория борьбы за существование. – М., Наука, 1976 .
2.
Понтрягин
Л.С., Андронов А.А., Витт А.А. ЖЭТФ. – 1933. – Т.3. – Вып.3. – С.165.
3.
Романовский Ю.М.,
Степанова Н.В., Чернавский Д.С. Математическое моделирование в
биофизике. – М.: Наука,1975.
4.
Свирежев
Ю.М. Нелинейные волны, диссипативные структуры и катастрофы в экологии. –
М.:Наука, 1987.
5.
Хорстхемке В.,
Лефевр Р. Индуцированные шумом переходы. Теория и применение в физике,
химии и биологии. – М.: Мир, 1987.
6.
Диментберг
М.Ф. Случайные процессы в динамических системах с переменными параметрами. –
М.: Наука, 1989.
7.
Климонтович
Ю.Л. Успехи физических наук. 164 (8). – 1994. – С. 813-842.
8.
Ершов
Ю.А. Журнал физической химии. – 1998. – Т.72. – №3. – С.553-559.
9.
Котин В.В.
Тезисы шестой международной конференции «Математика. Компьютер. Образование». –
М.: Прогресс-Традиция, 1999. – С. 145.
10.
Дьяконов
В.П. Математическая система MAPLE V R3/R4/R5. – М.: Солон, 1998.
11.
Mathcad
User’s Guide. Mathcad 8 Professional Academic. MathSoft, Inc., Cambridge, MA,
1998.